Prior diagnoses of severe infections are associated with a higher risk of neurodegenerative diseases and dementia. However, some of these associations did not remain statistically significant after adjusting for multiple comparisons, indicating that further research is needed to confirm these findings. Studies suggest acute bacterial and viral infections may be associated with brain volume loss. Reductions in total brain volume and grey matter thickness have been observed following infection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Besides, accelerated atrophy of white matter has been observed after symptomatic herpetic infections.
Systemic infections might influence dementia risk and neurodegeneration. Changes in circulating inflammatory proteins in response to immune challenges can impact brain health. Further, while select immunological markers have been linked to preceding inflammatory events and ensuing cognitive performance, how infections relate to diverse immunological proteins and which proteins predict atrophy-susceptible brain regions is unclear.
The study and findings
In the present study, researchers focused on identifying specific immunological proteins that predict changes in brain regions vulnerable to infection-related atrophy. They used data from the Baltimore Longitudinal Study of Aging and examined changes in brain volume between people with and without a history of any of fifteen select infections. Overall, 982 cognitively normal persons aged 65.4 years, on average, were included.
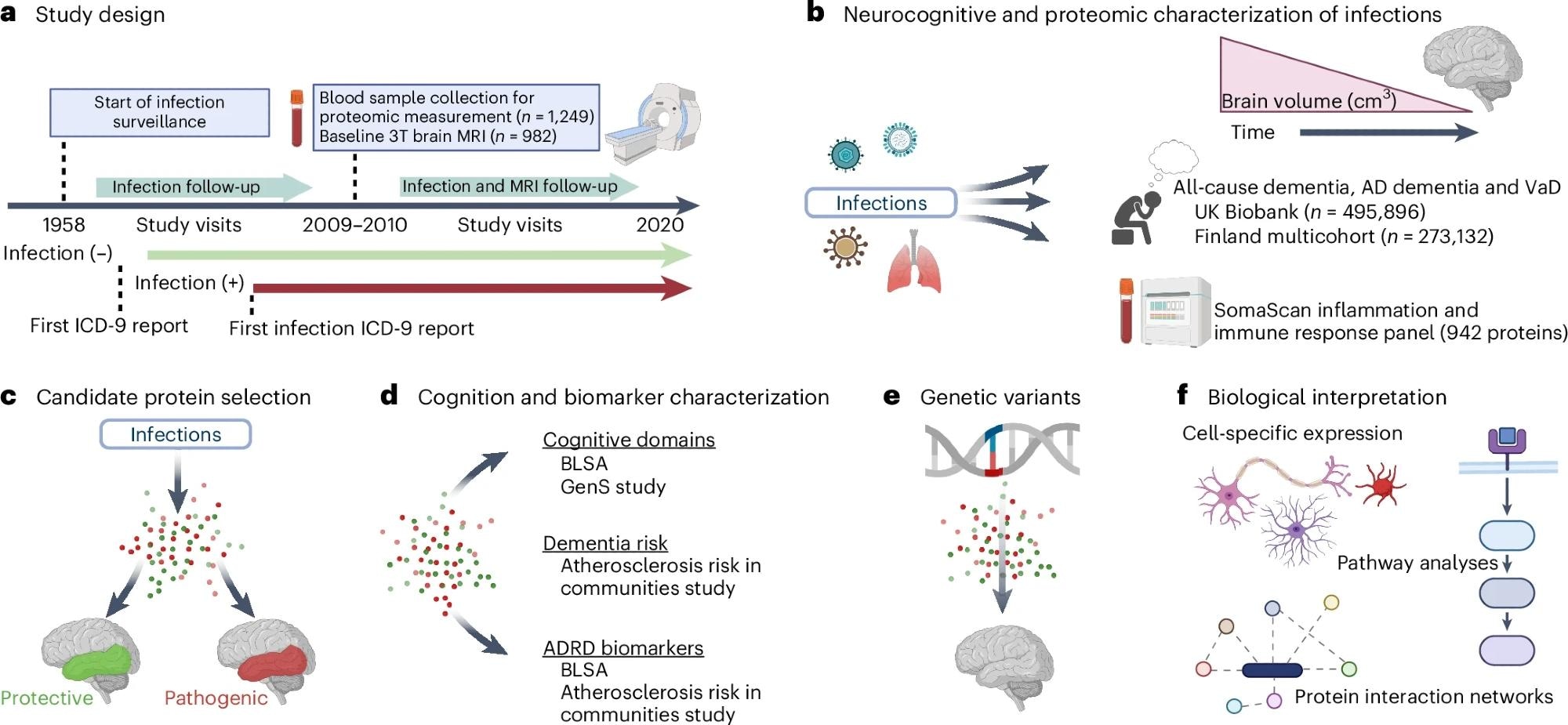 a, BLSA participants were classified according to the presence or absence of infection diagnoses using ICD-9 codes collected at study visits as early as 1958. Repeated 3T MRI scans were initiated in 2009–2010. Blood samples were collected at the initial 3T MRI scan and, for a subset of participants, at the time of a first PET scan as part of a separate study. b, Analyses examined how infection diagnoses are associated with brain volume changes over time and an immunological plasma proteome in the BLSA, as well as risk of all-cause dementia, AD dementia and VaD in the UK Biobank and a Finnish multicohort sample (the FPS study, the HeSSup study, and the STW study). c, Candidate proteins were selected if they were associated with an infection and related to changes in brain regions vulnerable to infection-specific atrophy, and were defined as protective or pathogenic, depending on whether they predicted preserved or reduced longitudinal brain volumes, respectively. d, Candidate proteins were related to longitudinal performance across five cognitive domains (the BLSA), cross-sectional performance across five cognitive domains (the GenS study), dementia risk (the ARIC study), and ADRD biomarkers (Aβ42/40, GFAP, NfL, pTau-181; the BLSA and the ARIC study). e, Genetic variants that influenced candidate protein levels were associated with changes in brain volumes in the BLSA and an external cohort (the ENIGMA consortium). f, The biological implications and functional relevance of candidate proteins were assessed using a variety of complementary analytical tools and open-source databases. All panels were created with BioRender.
a, BLSA participants were classified according to the presence or absence of infection diagnoses using ICD-9 codes collected at study visits as early as 1958. Repeated 3T MRI scans were initiated in 2009–2010. Blood samples were collected at the initial 3T MRI scan and, for a subset of participants, at the time of a first PET scan as part of a separate study. b, Analyses examined how infection diagnoses are associated with brain volume changes over time and an immunological plasma proteome in the BLSA, as well as risk of all-cause dementia, AD dementia and VaD in the UK Biobank and a Finnish multicohort sample (the FPS study, the HeSSup study, and the STW study). c, Candidate proteins were selected if they were associated with an infection and related to changes in brain regions vulnerable to infection-specific atrophy, and were defined as protective or pathogenic, depending on whether they predicted preserved or reduced longitudinal brain volumes, respectively. d, Candidate proteins were related to longitudinal performance across five cognitive domains (the BLSA), cross-sectional performance across five cognitive domains (the GenS study), dementia risk (the ARIC study), and ADRD biomarkers (Aβ42/40, GFAP, NfL, pTau-181; the BLSA and the ARIC study). e, Genetic variants that influenced candidate protein levels were associated with changes in brain volumes in the BLSA and an external cohort (the ENIGMA consortium). f, The biological implications and functional relevance of candidate proteins were assessed using a variety of complementary analytical tools and open-source databases. All panels were created with BioRender.
Of these, 67% were White, and 55.2% were females. About 43% of participants had no infection history, and over 10% had at least two infection diagnoses. The regions of interest (ROIs) were lobar, white matter, grey matter, total brain, and Alzheimer’s disease (AD) signature region volumes. If a significant association was found between infection and ROI, follow-up analyses on lobar grey/white matter were performed.
Six infections were associated with increased brain volume loss, especially in temporal grey and white matter regions. These were influenza, human herpes virus infections (HHVs), lower (LRTIs) and upper (URTIs) respiratory tract infections, skin and subcutaneous infections, and miscellaneous viral infections. It should be noted, however, that after applying a false discovery rate (FDR) correction, these associations were not statistically significant, suggesting that these findings should be interpreted with caution. Besides, follow-up analyses indicated that influenza-related brain volume loss was specific to occipital and temporal lobe grey matter.
Herpetic infection-related reductions in white matter volumes were specific to the temporal lobe. Accelerated atrophy related to miscellaneous viral infections was exclusive to grey matter. URTIs were associated with accelerated loss in the temporal lobe but were not specific to grey or white matter. LRTI-related temporal lobar volume loss was specific to white matter, while, in the occipital lobe, loss was observed in grey and white matter.
Grey matter atrophy associated with skin and subcutaneous infections was specific to occipital and temporal lobes. However, the overall history of infections or the number of infections did not show a statistically significant association with brain volume changes after corrections. The number of infections or the history of any infection was not associated with volume changes. Next, the team investigated associations between brain loss-related infections and the risk of all-cause, AD, and vascular (VaD) dementia using United Kingdom Biobank data. They found that the six infections were associated with a higher risk of all-cause dementia.
Five and four infections were associated with VaD and AD dementia, respectively. Particularly, the risk was most elevated for VaD, which aligns with previous findings that postinfection immune alterations may contribute more strongly to cerebrovascular pathology.
Next, the team assessed the plasma immune proteomic signature of brain atrophy-related infections. Influenza was found to be associated with the highest number of differentially expressed proteins (141), while miscellaneous viral infections were associated with the fewest (21).
Overall, 260 proteins were associated with at least one infection, and 38 were associated with multiple infections. Of the 260 proteins, 35 were associated with changes in the brain volume and were termed candidate proteins.
The (candidate) proteins were further defined as pathogenic or protective based on whether they reduced or preserved brain volumes respectively. Protective proteins were reduced among individuals with infection history, while pathogenic proteins were increased. Further, the researchers examined whether candidate proteins were associated with cognitive performance.
Ten protective proteins were associated with preserved verbal memory performance, and some were associated with preserved verbal fluency or visuospatial performance. Four pathogenic proteins were associated with rapid performance reductions. Interestingly, one pathogenic protein (CD27) was unexpectedly linked to preserved visuospatial abilities, highlighting the complexity of these associations.
Next, the team evaluated whether candidate proteins are related to AD and neurodegeneration plasma biomarkers.
Twelve protective proteins that decreased post-infection were associated with a significantly elevated amyloid beta (Aβ)42/40 ratio, and 11 were associated with phosphorylated Tau (pTau)-181. One pathogenic protein was associated with increased neurofilament light chain (NfL) and lower Aβ42/40 ratio, and several others had negative associations with pTau-181 and NfL.
Lastly, the team investigated whether genetic variants affecting candidate protein levels relate to brain volume changes. They found that genetic variants linked to lower pathogenic protein levels were associated with preserved brain volumes, even in regions susceptible to atrophy. However, not all genetic variants were protective; some that reduced pathogenic protein levels were paradoxically associated with accelerated volume loss, indicating a complex relationship.
Conclusions
The study revealed how infections, such as LRTIs, URTIs, influenza, HHVs, skin and subcutaneous infections, and miscellaneous viral infections, are associated with elevated brain atrophy and future dementia risk. While some associations did not hold up after multiple testing corrections, the overall pattern suggests a link between infections and neurodegeneration. Thirty-five immunologically relevant plasma proteins differentially expressed after infection were identified.
These proteins were related to changes in brain regions vulnerable to infection-specific atrophy. Some proteins were linked to cognitive decline and plasma biomarkers of neurodegeneration and AD pathology. Together, the findings support the role of infection in dementia risk and reveal the molecular mediators by which infections contribute to neurodegeneration. Further research is needed to fully understand these complex relationships and their implications for dementia risk.